This chapter provides an overview of basic and essential neuroanatomy and neurophysiology as a quick reference to assist the reader in understanding the underlying principles of neurological function and dysfunction as a basis for nursing management. Further discussion of anatomy and physiology is included in many other chapters to enhance understanding. Neuroanatomy and neurophysiology texts should be consulted if more detail is desired.
EMBRYONIC DEVELOPMENT OF THE NERVOUS SYSTEM
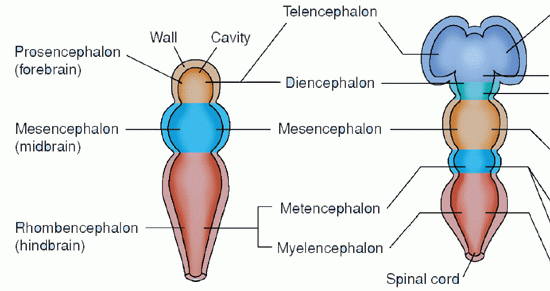
The human brain is composed of about 100 billion cells. The nervous system is one of the first recognizable features in embryonic development. From a simple longitudinal invagination on the dorsal portion of the ectodermal layer, a neural groove and neural tube form at about 3 weeks. At the cranial end of the neural tube, rapid and unequal growth occurs, giving rise to the three primary vesicles of the brain: the prosencephalon, or forebrain; the mesencephalon, or midbrain; and the rhombencephalon, or hindbrain. Early in the second fetal month, these vesicles, in turn, become five cerebral areas: the telencephalon, or endbrain; the diencephalon, or between brain; the mesencephalon; the metencephalon, or afterbrain; and the myelencephalon, or spinal brain (Table 5-1). By 7 weeks, the brain and spinal cord are apparent. At 12 weeks, the brain is the size of a large pea.
Concurrently, cells of the neural tube form two types of cells: spongioblasts, which give rise to the neuroglia (glia) cells, and neuroblasts, which give rise to the nerve cells (neurons). Processes from the neuroblasts form the white matter of the brain. Some of these processes leave the brain and spinal cord to form the fibers of the cranial and ventral roots of the spinal nerves.
Throughout the prenatal period, there is further growth and refinement of the nervous system. All the neurons that a person will ever have are present at birth. These highly specialized cells do not have mitotic capacity and, therefore, are not replaceable. At birth, the brain is about one quarter the size of an adult brain. Abnormalities in neural tube development can result in spinal bifida, meningocele, or myelomeningocele.
CELLS OF THE NERVOUS SYSTEM
From the ectodermal layer, two types of cells develop: neurons and neuroglia cells. Neurons are the basic anatomic and functional unit of the nervous system. Neuroglia cells provide a variety of supportive functions for the neurons.
Neuroglia Cells
The term glia comes from a Greek word meaning “glue” or “holding together.” In this regard, the glia cells provide structural support, nourishment, and protection for the neurons of the nervous system. There are 5 to 10 times more neuroglia cells than there are neurons. About 40% of the brain and spinal cord is composed of neuroglia cells.
From a clinical viewpoint, neuroglia cells are important because they can divide by mitosis and are the major source of primary tumors of the nervous system. In the central nervous system (CNS), glia is subdivided into four main types: astrocytes, oligodendrocytes, ependymal cells, and microglia. In the peripheral nervous system (PNS), Schwann cells form myelin sheaths.
TABLE 5-1 DEVELOPMENT OF THE PRIMARY VESICLES
PRIMARY VESICLES
SUBDIVISIONS
STRUCTURES THAT ARISE
VENTRICULAR SYSTEM
Cerebral hemisphere, corpus callosum, basal ganglia, olfactory tracts
Lateral ventricles and part of third ventricle
Thalamus
Hypothalamus
Most of third ventricle
Midbrain
Cerebral aqueducts
Pons
Cerebellum
Fourth ventricle
Medulla oblongata
Fourth ventricle and part of central canal
Astrocytes have multiple processes extending from the cell body that give it a star-like appearance. Some astrocytic processes may terminate as swellings called end-feet on neurons and blood vessels. Functions attributed to astrocytes include providing nutrition for neurons, regulating synaptic connectivity, removing cellular debris, and controlling movement of molecules from blood to brain (part of the blood-brain barrier [BBB]).
On microscopic examination, oligodendrocytes have few branching processes. Oligodendrocytes produce the myelin sheath of the axonal projections of neurons in the CNS. An individual cell can maintain the myelin sheaths of several axons.
Ciliated ependyma cells line the ventricular system and the choroid plexuses. They aid in the production of cerebrospinal fluid (CSF) and act as a barrier to foreign substances within the ventricles, preventing them from entering cerebral tissue.
Microglia cells are minute cells that are scattered throughout the CNS and have a phagocytic function. They remove and disintegrate the waste products of neurons.
Schwann cells function similarly to oligodendrocytes, forming the insulating myelin sheaths around axons to facilitate saltatory conduction of impulses in the PNS.
Neurons
Neurons vary from 5 to 100 µm in diameter. As the basic anatomic and functional unit in the nervous system, the neuron has a number of functions: responding to sensory and chemical stimuli, conducting impulses, and releasing specific chemical regulators.
Neurons are classified as unipolar, bipolar, or multipolar. Unipolar neurons possess only one process or pole. This process divides close to the cell body. One branch, called the peripheral process, carries impulses from the periphery toward the cell body. The other branch, called the central process, conducts the impulse toward the spinal cord or the brainstem. Bipolar neurons are found only in the spinal and vestibular ganglia, the olfactory mucous membrane, and one layer of the retina. The anatomic structure is peculiar to the organ in which it is found. Most neurons in the nervous system are multipolar. These neurons consist of a cell body, one long projection (the axon), and one or more shorter branches (the dendrites).
There are three major components of a neuron: a cell body, which constitutes the main part of the neuron; a single axon, or axis cylinder, which consists of a long projection extending from the cell body; and several dendrites, which are thin projections extending from the cell body into the immediate surrounding area. The axon carries impulses away from the cell body whereas the dendrites direct impulses toward the cell body.
COMPONENTS OF THE CELL BODY
The main organelles of the neuronal cell body include the nucleus, the cell membrane, and the cytoplasm. There are organelles within each of these structures that are important and are mentioned here briefly.
The nucleus is a double-membrane structure that contains chromatin and a prominent nucleolus. Chromatin is the threadlike structures in the cell nucleus that consist primarily of deoxyribonucleic acid (DNA) and protein. DNA contains genes and the genetic code or information about the cell. The nucleolus contains ribonucleic acid (RNA). RNA is the “messenger” from the genes of the nucleus; it contains the code for synthesis of specific cellular proteins.
A lipid bilayer is the basic cell membrane structure. The cell membrane is a solution of globular proteins dispersed in a fluid phospholipids matrix. The cell membrane creates the parameters of the cell body, enclosing the cytoplasm within its border. The main purpose of the cell membrane is to control the interchange of material between the cell and its environment.
The cytoplasm contains smooth and rough endoplasmic reticula, Nissl bodies, Golgi apparatus, mitochondria, lysosomes, neurotubules, and neurofibrils. The endoplasmic reticulum of the cytoplasm is a network of tubular membranous structures. There are two types of endoplasmic reticulum, smooth and rough. The smooth endoplasmic reticulum serves as the site for enzyme reactions. Centrioles are found in the cytoplasm and take part in cell division. Nissl bodies are masses of granular (rough) endoplasmic reticulum with ribosomes, which are the protein-synthesizing machinery of the neuron. The endoplasmic reticulum system connects with the nucleus at that portion of the reticulum called the Golgi apparatus. Substances formed in different parts of the cell are transported throughout the cell by means of this system. The Golgi apparatus provides for two interrelated functions: further modification of protein by adding carbohydrates and temporary storage and separation of protein types, depending on their function and destination. It is also responsible for the formation of substances important for the digestion of intracellular material.
Mitochondria are structures that serve as the site for production of most cellular energy. Cell nutrients are oxidized to produce carbon dioxide and water. The energy released is used to produce adenosine triphosphate (ATP). Lysosomes isolate the digestive enzymes of a cell from the cytoplasm to prevent cell destruction. They are involved in digestion of phagocytosis products and worn-out organelles.
The elongated axons or dendrites can extend 1 m or more from the cell body. These fibers require protein and other substances produced in the cell body that must be transported from the cytoplasm by a process called axoplasmic flow.Neurotubules carry out part of axoplasmic transport. Neurofibrils are delicate thread-like structures within the cytoplasm and the axon hillock that assist in the transport of cellular material.
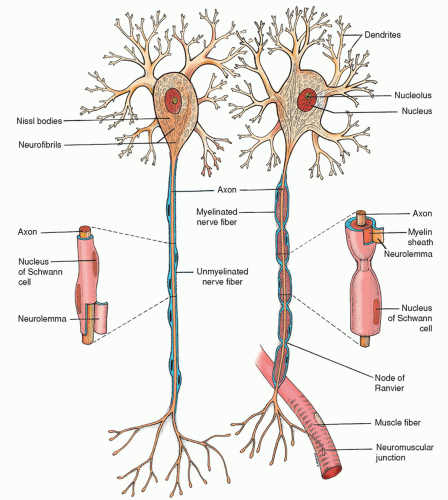
Figure 5-1 ▪ Typical neurons: unmyelinated fiber (left); myelinated fiber (right). (From Chaffee, E. E., & Lytle, I. M. (1980). Basic physiology and anatomy. Philadelphia: J. B. Lippincott.)
CELL PROCESSES: AXONS AND DENDRITES
Axons and dendrites constitute the cell processes. Dendrites usually extend only a short distance from the cell body and branch profusely. By contrast, an axon can extend for long distances from the cell body before branching near the end of the projection.
Many axons in the PNS are covered by a myelin sheath composed of a white, lipid substance that acts as an insulator for the conduction of impulses. Nerve fibers enclosed in such a sheath are referred to as myelinated; those without the myelin sheath are referred to as unmyelinated (Fig. 5-1). As a rule, larger neuron fibers are myelinated whereas smaller fibers are unmyelinated.
The myelin sheath is formed by Schwann cells that encircle the axons. When several Schwann cells are wrapped around an axon, their outer layer (sheath of Schwann) encloses the myelin sheath. This outer layer is called the neurolemma and is said to be necessary for the regeneration of axons. The myelin sheath itself is a segmented, discontinuous layer that is interrupted at intervals by the nodes of Ranvier. The distance from one node to the next is called an internode. Each internode is formed by, and surrounded by, one Schwann cell. At the junction between each of the two successive Schwann cells along the axon, a small noninsulated area remains where ions can easily flow between the extracellular fluid and the axon. It is this area that is known as the node of Ranvier. In the CNS, the oligodendroglial cells provide the myelination of the neurons, similar to the role of the Schwann cells in the PNS. Myelination abnormalities can result in dysfunction in the CNS such as multiple sclerosis or PNS such as Guillain-Barré syndrome.
PHYSIOLOGY OF NERVE IMPULSES
Resting Membrane Potential of the Neurons
Although a resting neuron is not conducting an impulse, it is considered to be a charged cell. The difference in electrical charge on either side of the membrane is called the potential difference and is related to the unequal distribution of potassium and sodium on either side of the membrane. The neuronal cell membrane is electrically polarized to a resting membrane potential of about -80 mV. By convention, the outside of the membrane is considered to be at 0 mV; therefore, at a resting potential of -80 mV means the inside of the membrane is 80 mV negative to the outside. The resting membrane potential results from the separation of charges across the cell membrane.
The cell membrane is both semipermeable and selectively permeable. The area outside the cell is called the interstitial space; the area inside the cell is called the intracellular space. Sodium ions (Na+) and chloride ions (Cl−) are found in much greater concentrations in the interstitial space than in the cell. The potassium ion (K+) and organic protein material are found in high concentrations within the cell. Diffusion is responsible for the movement of Na+ and K+ to the other side of the membrane. If they ever reach equilibrium there will no longer be a resting membrane potential. The maintenance of the resting membrane potential lies with K+ and Na+. Two transport proteins are primarily responsible for the resting membrane potential: a K+ leak channel that permits K+ to diffuse out of the cell, and the sodium potassium pump (Na+, K+-ATPase). The Na+, K+-ATPase pumps two K+ back into the cell at the same time that it pumps three Na+ out of the cell. Both processes keep the concentration gradient across the cell membrane constant. The concentration of dissolved ions in a solution is a potential source of energy to drive cellular processes.
Action Potential of the Neuron
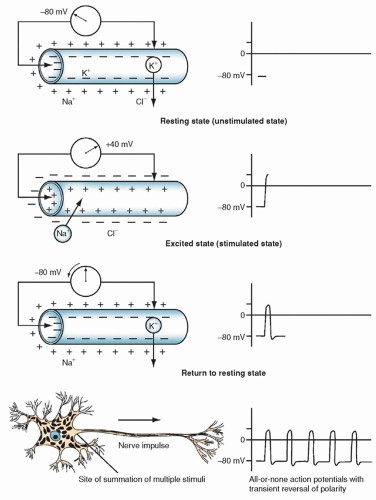
The fluid and ions in the intracellular space create a highly conductive solution. The large diameter (10 to 80 µm) allows for unrestricted conduction of impulses from one part of the interior of the cell to the other. Various stimuli can change the permeability of the cell membrane to certain ions, resulting in alterations in the membrane potential (Fig. 5-2). The stimuli must be of sufficient magnitude to conduct an impulse and thus create an action potential, which is the fundamental unit of signaling in the nervous system.
Many simultaneous discharges at the synaptic junction must occur to create a sufficient effect on the cell membrane. The membrane potential reverses, and the intracellular surface becomes positive (approximately +20 to +40 mV). When the action potential is realized, there is a sudden reversal of the sodium and potassium relationship across the cell membrane of the axon. This event is called depolarization. The neuron receives an influx of sodium and loses potassium to the interstitial space; the time required is only a few milliseconds. With the change in polarity, an impulse is conducted from one neuron to the next at the same amplitude and speed. The cell repolarizes and returns to its resting membrane potential. This sequence of events occurs during the conduction of an impulse in an unmyelinated nerve.
Saltatory Conduction
In myelinated nerves, an action potential hops from one node of Ranvier to the next as a means of rapidly conducting an impulse. This is called saltatory conduction. Although ions cannot flow out through the myelin sheath of myelinated nerves, the break in the myelin sheath at the nodes of Ranvier (Fig. 5-1) provides a perfect route of escape. At this point, the membrane is several times more permeable than many unmyelinated nerves. Impulses are conducted from node to node rather than continuing along the entire span of the axon, as is the case in unmyelinated nerves. Saltatory conduction is advantageous because it increases the velocity of an impulse and conserves energy (because only the nodes depolarize). The velocity of an impulse depends on both the thickness of the myelin and the distance between the internodes. As these two factors increase, the velocity of the impulse also increases.
Synapse
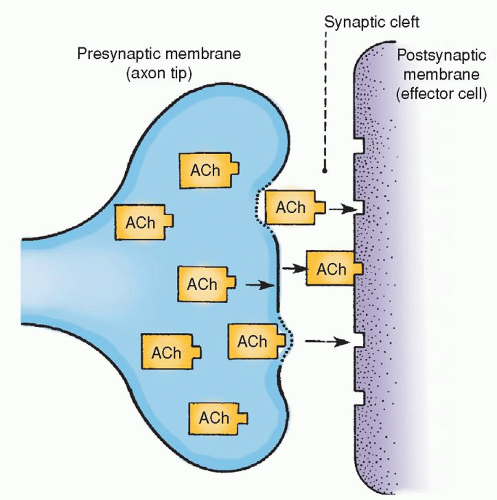
The junction between one neuron and the next at which an impulse is transmitted is called the synaps. There are three anatomic structures that are necessary for an impulse to be transmitted at a synapse. These include the presynaptic terminals, the synaptic cleft, and the postsynaptic membrane (Fig. 5-3).
Presynaptic terminals are either excitatory or inhibitory. An excitatory presynaptic terminal secretes an excitatory substance into the synaptic cleft, thereby exciting the effector neuron. The inhibitory presynaptic terminal secretes an inhibitory transmitter that, when secreted into the synaptic cleft, inhibits the effector neuron. The excitatory or inhibitory transmitter secretions arise from the synaptic vesicle in the presynaptic terminal of the axon. Mitochondria in the axon supply the ATP to synthesize new transmitter secretions. The synaptic cleft is the microscopic space (200 to 300 Å) between the presynaptic terminal and the receptor area of the effector cell. The postsynaptic membrane is that part of the effector membrane distal to the presynaptic terminal.
When an action potential spreads over the presynaptic terminal, the membrane depolarizes, emptying some of the contents of the presynaptic vesicles into the synaptic cleft. The released transmitter changes the permeability of the postsynaptic membrane. This results in either excitation or inhibition of the neuron, depending on the type of transmitter substances secreted into the synaptic cleft.
Neurotransmitters
Chemical substances found in the CNS that excite, inhibit, or modify the response of another cerebral cell or cells are called neurotransmitters. The presynaptic terminals of one neuron release the chemical that affects particular postsynaptic cells of another neuron. Generally, each neuron releases the same transmitter at all of its separate terminals. Over 100 neurotransmitters have been identified. They include specific amines (acetylcholine, serotonin), catecholamines (dopamine, epinephrine, and norepinephrine), amino acids (γ-amino butyric acid [GABA], glutei acid, lysine, and substance P), and polypeptides (endorphins and enkephalins). Table 5-2 summarizes the major neurotransmitters.
Figure 5-2 ▪ The ionic and electrical changes that occur in a neuron when stimulated. (From Snell, R. S. (1997). Clinical neuroanatomy for medical students (4th ed.). Philadelphia: Lippincott Williams & Wilkins.)
Postsynaptic Membrane—Excitation and Inhibition
The postsynaptic membrane (the dendrite cell body region of the effector neuron) initiates its response to stimuli by decremental conduction through the synapse; that is, the impulse becomes progressively weaker during more prolonged periods of excitation. Stimulation of the effector cell at the dendrite cell body can create an action potential. For the action potential to be fired, however, the intensity of an impulse must be sufficient to fire the initial segment of axon (just distal to the axon hillock), where the action potential is initiated. It is said that any factor that increases the potential inside the cell body at any given point also increases the potential throughout the cell body.
Because of differences in the cell membrane and shape of the cell, the intracellular voltage necessary to elicit an action potential will vary at different points on the cell membrane. The most sensitive point is the initial segment of the axon, but the impulse must be of sufficient magnitude to depolarize the axon. Excitation is the response of the postsynaptic membrane to the neurotransmitter substance that lowers the membrane potential to form an excitatory postsynaptic potential (EPSP). The potential is a small depolarization that conducts itself by decrement. The cell membrane is made more permeable to sodium, potassium, and chloride. Sodium ions rush into the neuron whereas potassium ions leave the cell through the postsynaptic membrane.
Inhibition acts on a cell so that it is more difficult for it to fire. The inhibitory neurotransmitters increase permeability to only potassium and chloride ions at the synaptic membrane. The membrane potential is raised to form the inhibitory postsynaptic potential (IPSP).
Presynaptic Inhibition
Another type of inhibition, presynaptic inhibition, results from inhibitory knobs being activated on the presynaptic terminal fibrils and synaptic knobs of an axon. When the inhibitory knobs are activated, they secrete a neurotransmitter substance that partially depolarizes the terminal fibrils and excitatory synaptic knobs. As a result, the velocity of the action potential that occurs at the membrane of the excitatory knob is depressed. This action greatly reduces the quantity of excitatory neurotransmitter released by the knob and suppresses the degree of excitation of the neuron.
Figure 5-3 ▪ Conduction at synapses. Diagram shows the release of a neurotransmitter (in this case acetylcholine [Ach]) from the presynaptic membrane (axon tip) into the synaptic cleft, where it bonds to the receptor sites of the postsynaptic membrane (effector cell). (From DeMyer, W. (1998). Neuroanatomy (2nd ed.). Baltimore: Williams & Wilkins.)
FUNCTIONS AND DIVISIONS OF THE NERVOUS SYSTEM
The nervous system controls the motor, sensory, autonomic, cognitive, and behavioral functions of the body. It is divided into a hierarchy with three major functional units.
Spinal cord level—the lowest functional level; controls automatic motor responses, such as reflexes.
Brainstem and subcortical level—the second functional level; controls blood pressure, respirations, equilibrium, and primitive emotions.
Cortical level—the highest level; responsible for cognition (storage of information, thinking, memory, and abstraction).
The nervous system is also divided into the CNS and the PNS. The CNS is composed of the brain and the spinal cord. The PNS includes the 12 pairs of cranial nerves (CNs), the 31 pairs of spinal nerves, and the autonomic nervous system (ANS), which subdivides into the sympathetic and parasympathetic nervous systems.
CRANIAL AND SPINAL BONES
The purpose of the skull and vertebral column is to protect the most vulnerable parts of the nervous system—the brain and the spinal cord.
Skull
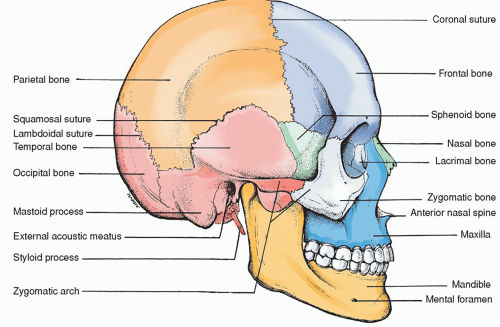
The skull is the bony framework of the head; it is composed of the 8 bones of the cranium and the 14 bones of the face. Knowledge of the anatomy of both the external and internal surfaces of the bones (Figs. 5-4 and 5-5) is helpful in understanding the pathophysiology of craniocerebral trauma.
The cranium is defined as that part of the skull that encloses the brain and provides a protective vault for this vital organ. The bones that compose the cranium are the frontal, occipital, sphenoid, and ethmoid, as well as the two parietal and temporal bones.
The frontal bone forms the forehead and the front (anterior) part of the top of the skull. The supraorbital arches form the roofs of the two orbits. Frontal sinuses are also located in this bone. The inner table of the frontal bone has a highly irregular bony surface.
The occipital bone is the large bone at the back (posterior) of the skull that curves into the base of the skull. The significant markings include the large hole (foramen magnum) in the base of the skull and also the occipital condyles located on either side of the foramen magnum that fit into depressions on the first cervical vertebra.
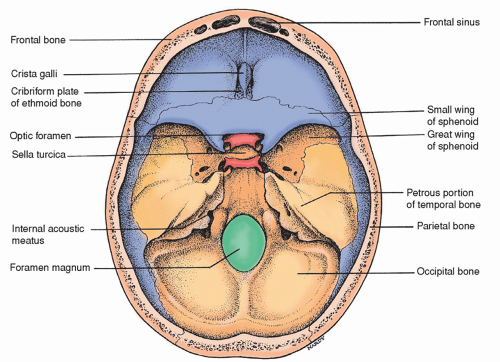
The sphenoid bone is a wedge-shaped bone thought to resemble a bat’s wings. The significant bone markings include a body, lesser wings, greater wings, the pterygoid process, the sella turcica (Latin for “Turk’s saddle”), and the clivus. The clivus is the slanted dorsal surface of the body of the sphenoid bone between the sella turcica and basilar process of the sphenoid bone. The hypophysis (pituitary gland) is located in the region of the sella turcica.
The ethmoid bone, largely hidden between the two orbits, contains both perpendicular and horizontal plates as well as two lateral masses. The horizontal plate, also called the cribriform plate, forms part of the base of the skull through which the olfactory nerves (first CNs) travel. The perpendicular plate forms part of the nasal septum whereas the lateral masses are part of the ethmoid sinuses.
The temporal bones are situated at the sides and base of the skull and consist of three anatomic divisions: the squamous, mastoid, and petrous portions. The temporal bone is highly irregular, both on the internal and external surfaces.
The squamous portion is very thin just above the auditory meatus. It contains the zygomatic process externally; internally, there are numerous eminences and depressions to accommodate the contour of the cerebrum. Two well-marked internal grooves are evident for the branches of the middle meningeal artery, a common point of trauma with head injury.
The mastoid portion is perforated by many foramina, including a larger foramen, the mastoid foramen, which contains a vein that drains the lateral sinus and a small artery to supply the dura mater. The mastoid process is also contained in this portion of the temporal bone.
The petrous portion is so named because it is extremely dense and stone-like in its hardness. There is a pyramidal process that is directed inward and is wedged at the base of the skull between the sphenoid and occipital bones. The internal plate of the petrous portion of the temporal bone is proximal to the branches of the middle meningeal artery.
The parietal bones, which fuse on the top of the skull, form the sides of the skull. Externally, they are smooth and convex; internally, the surface is concave with some depressions to accommodate the convolutions of the cerebrum and the grooves for the middle meningeal artery. With these exceptions, the bone has a regular inner surface.
TABLE 5-2 NEUROTRANSMITTERS: SOURCE AND ACTION
NAME
SECRETION SOURCE
ACTION
Amines
Acetylcholine (ACh)
First neurotransmitter identified
Neurons in many areas of brain
Large pyramidal cells (motor cortex)
Usually excitatory
Inhibitory effect on some of parasympathetic nervous system (e.g., heart by vagus)
Chief transmitter of parasympathetic nervous system
Some cells of basal ganglia
Motor neurons that innervate skeletal muscles
Preganglionic neurons of autonomic NS
Postganglionic neurons of parasympathetic NS
Postganglionic neurons of sympathetic NS
Serotonin (5-HT)
Controls body heat, hunger, behavior, and sleep
Nuclei originating in the median raphe of brainstem and projecting to many areas (especially the dorsal horns of the spinal cord and hypothalamus)
Inhibitor of pain pathway cord; helps to control mood and sleep
Catecholamines
Dopamine (DA)
Affects control of behavior and fine movement
Neurons on the substantia nigra; many neurons of the substantia nigra send fibers to the basal ganglia that are involved in coordination of skeletal muscle activity
Usually inhibitory
Norepinephrine (NE)
Chief transmitter of sympathetic nervous system
Many neurons whose cell bodies are located:
In brainstem and hypothalamus (controlling overall activity and mood)
Most postganglionic neurons of sympathetic NS
Usually excitatory, although sometimes inhibitory
Some excitatory and some inhibitory
Amino Acids
γ-aminobutyric acid (GABA)
Nerve terminals of the spinal cord, cerebellum, basal ganglia, and some cortical areas
Excitatory
Glutamic acid
Presynaptic terminals in many sensory pathways; cerebellum mossy fibers
Excitatory
Glycine
Synapses in spinal cord
Inhibitory
Substance P
Pain fiber terminals in the dorsal horns of the spinal cord; also, the basal ganglia and hypothalamus
Excitatory
Polypeptides
Enkephalin
Nerve terminals in the spinal cord, brainstem, thalamus, and hypothalamus
Excitatory to systems that inhibit pain; binds to the same receptors in the CNS that bind opiate drugs
Endorphin
Pituitary gland and areas of the brain
Binds to opiate receptors in the brain and pituitary gland; excitatory to systems that inhibit pain
NS, nervous system; CNS, central nervous system.
SUTURES OF THE SKULL
The bones of the skull join at various places, known as suture lines. The four major sutures of the skull include the following (Fig. 5-4).
Sagittal suture: the midline suture formed by the two parietal bones joining on the top of the skull.
Coronal suture (frontoparietal): connecting the frontal and parietal bones transversely.
Lambdoidal suture (occipitoparietal): connecting the occipital and parietal bones.
Basilar suture: created by the junction of the basilar surface of the occipital bone with the posterior surface of the sphenoid bone.
SPINE
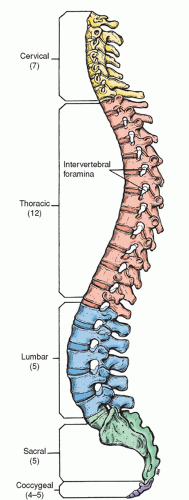
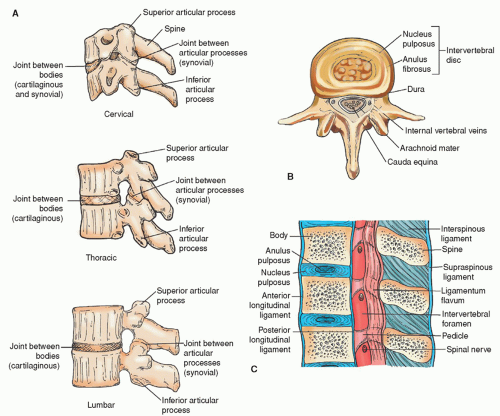
The spine is a flexible column formed by a series of bones called vertebrae, each stacked one on another to support the head and trunk. The vertebral column is made up of 33 vertebrae: 7 cervical vertebrae, 12 thoracic or dorsal vertebrae, 5 lumbar vertebrae, 5 sacral vertebrae (fused into one), and 4 coccygeal vertebrae (fused into one) (Fig. 5-6). Each vertebra consists of two essential parts, an anterior solid segment or body, and a posterior segment or arch. Two pedicles and two laminae supporting seven processes (four articular, two transverse, and one spinous) make up the arch (Fig. 5-7).
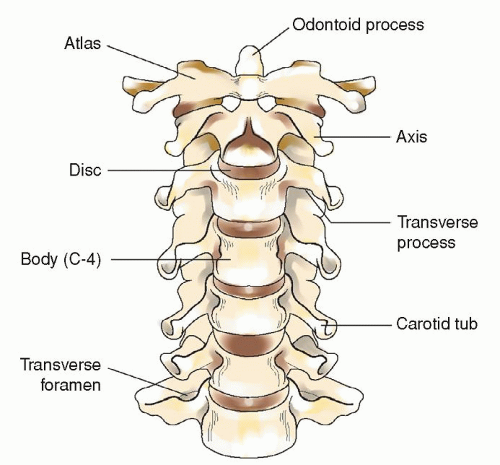
The cervical vertebrae are smaller than those in any other region of the spine. The first cervical vertebra is called the atlas whereas the second cervical vertebra is known as the axis. Each of these two vertebrae has a unique appearance. The axis has a perpendicular projection called the odontoid process on which the atlas sits (Fig. 5-8). The thoracic or dorsal vertebrae are intermediate in size, becoming larger as they descend the vertebral column. The lumbar vertebrae are the largest segments in the spine (Fig. 5-7).
Figure 5-4 ▪ Lateral view of the skull.
Figure 5-5 ▪ View of the base of the skull from above, showing the internal surfaces of some of the cranial bones.
Figure 5-6 ▪ Lateral view of the adult vertebral column.
The vertebral bodies are the largest part of the vertebrae, above and below which flattened surfaces are found for attachment of fibrocartilage. There are apertures for spinal nerves, veins, and arteries. The vertebrae are connected by means of the articular processes and the intervertebral fibrocartilage.
The arch of the vertebrae is composed of two pedicles, two laminae, a spinous process, four articular processes, and two transverse processes. The two pedicles are short, thick pieces of bone. The concavity above and below the pedicles creates the intervertebral notches from which the spinal nerves emanate. The two laminae are broad plates of bone. They complete the neural arch by fusing in the midline and enclose the spinal foramen, which protects the spinal cord. The upper and lower borders are rough in order to allow for the attachment of the ligamenta subflava. The spinous process projects backward from the laminae and serves as the attachment for muscles and ligaments. The four articular processes (two on either side) provide stability for the spine. The two transverse processes provide stability for the spine and serve as points of attachment for muscles and ligaments.
LIGAMENTS OF THE SPINE
The most important ligaments of the vertebral column are the anterior and posterior longitudinal ligaments and the ligamenta flava (Fig. 5-7). The anterior longitudinal ligament consists of longitudinal fibers firmly attached to the anterior surface of the vertebral bodies and intervertebral discs. The posterior longitudinal ligament is attached to the posterior surface of the vertebral bodies within the spinal canal. The ligamentum flavum consists of yellow elastic fibers that connect the laminae of adjacent vertebrae. The attachment pattern is unique in that the attachment is from the lower margin of the anterior surface of the superior lamina to the posterior surface of the upper margin of the inferior lamina.
The supraspinous ligament joins the spinous process tips from C7 to the sacrum. The interspinous ligaments connect adjacent spinous processes from their tips to their roots. The interspinals fuse with the supraspinals posteriorly and with the ligamentum flavum anteriorly. Such an arrangement controls vertebral movement to prevent excessive flexion. If violent force in any direction occurs, these ligaments can be ruptured, possibly causing injury to the vertebrae and spinal cord (see Chapters 18 and 19).
INTERVERTEBRAL DISCS
The intervertebral discs are fibrocartilaginous disc-shaped structures located between the vertebral bodies from the second cervical vertebra to the sacrum. They vary in size, thickness, and shape at different levels of the spine. The purpose of the intervertebral disc is to cushion movement. The central core, the nucleus pulposus, is surrounded by a fibrous capsule called the annulus fibrosus. As a result of aging and trauma, discs lose their water content and the tissue is more prone to injury.
MENINGES
Meninges cover both the brain and the spinal cord. The layers, from the outermost layer inward, are called the dura mater, the arachnoid, and the pia mater (Fig. 5-9).
The dura mater is a double-layer, whitish, inelastic, fibrous membrane that lines the interior of the skull. The outer layer of the dura is actually the periosteum of the bone. The inner layer is the thick membrane that extends throughout the skull and creates compartments. The dura lines various foramina that exit at the base of the skull. Sheaths for the nerves passing through these foramina are also formed by the dura.
Four folds of dura (Fig. 5-10) are situated within the skull cavity to support and protect the brain. They include the following.
Falx cerebri, a double fold of dura, descends vertically into the longitudinal fissure between the two hemispheres of the brain and partially divides the frontal lobe into a left and right side.
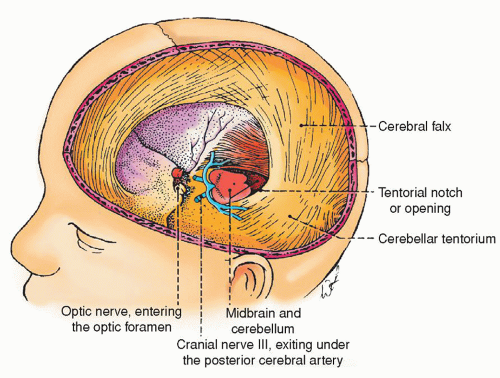
Tentorium cerebelli is a tent-like double fold of dura that covers the upper surface of the cerebellum, supports the occipital lobes, and prevents them from pressing on the cerebellum. The falx cerebri attaches midline to the tentorium. (The tentorium is an important anatomic point to note. The area above the tentorium is termed supratentorial whereas the area below it is called infratentorial. The nursing care given differs based on these two classifications, as discussed in Chapter 16.) In addition, the opening in the tentorium from which the brainstem emerges is called the tentorial notch. Herniation through this opening is called uncal herniation.
Falx cerebelli is found between the two lateral lobes of the cerebellum.
Diaphragma sella is a horizontal process that forms a small circular fold, thus creating a roof for the sella turcica.
Figure 5-7 ▪ A: Cervical, thoracic, and lumbar vertebrae. B: Third lumbar vertebra seen from above, showing the relationship between the intervertebral disc and the cauda equina. C: Sagittal section through three lumbar vertebrae, showing the ligaments and the intervertebral discs.
Figure 5-8 ▪ The cervical spine. Note odontoid process of C2 and the atlas, C1, positioned on top of C2.
The spinal dura is a continuation of the inner layer of the cerebral dura. The outer layer of the dura terminates at the foramen magnum, where it is replaced by the periosteal lining of the vertebral canal. The spinal dura encases the spinal roots, spinal ganglia, and spinal nerves. The spinal dural sac terminates at the second or third sacral level.
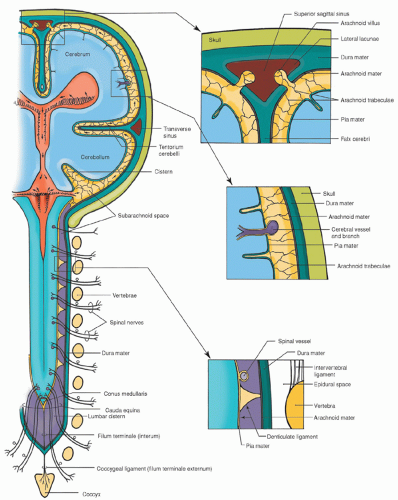
Figure 5-9 ▪ The central nervous system and its associated meninges. The top right box shows the superior sagittal sinus and arachnoid villi. Arachnoid villi remove cerebrospinal fluid (CSF) from the ventricles and deposit the CSF into the venous circulation. The middle right box shows the layers of tissue from the skull through the three layers of the meninges. The bottom right box shows the layers of spinal meninges. (From Haines, D. E. (2000). Neuroanatomy: An atlas of structures, sections, and systems (5th ed.). Philadelphia: Lippincott Williams & Wilkins.)
Figure 5-10 ▪ Cranial dura mater folds. With the skull and cerebral hemisphere removed, the falx cerebri and the tentorium cerebelli are shown. Note that cranial nerve III (oculomotor nerve) exits under the posterior cerebral artery. With lateral transtentorial herniation, the oculomotor nerve may be caught between the tentorium and posterior cerebral artery, resulting in dilation of the pupil. (From DeMyer, W. (1998). Neuroanatomy (2nd ed.). Baltimore: Williams & Wilkins.)
The second meningeal layer, the arachnoid membrane, is an extremely thin, delicate layer that loosely encloses the brain. The subdural space separates the dura mater from the arachnoid layer. Bleeding within this space (subdural hemorrhage) can occur with head injury. The subarachnoid space is not really a clear space because there is much spongy, delicate connective tissue between the arachnoid and pia mater layers. CSF flows in the subarachnoid space. The cisterna magnum is a space between the hemispheres of the cerebellum and the medulla oblongata. The arachnoid layer of the spinal meninges is a continuation of the cerebral arachnoid (Fig. 5-9). The arachnoid is a delicate, gossamer network of fine, elastic, fibrous tissue; it also contains blood vessels of varying sizes, which may be damaged by lumbar or cisternal puncture, resulting in hemorrhage.
The innermost layer of the meninges is called the pia mater. It is a mesh-like, vascular membrane that derives its blood supply from the internal carotid and vertebral arteries. The pia mater covers the entire surface of the brain, dipping down between the convolutions of the surface. Because the pia covers the gray matter, vascularity increases and minute perpendicular vessels extend for some distance into the cerebrum. The pia mater of the spinal cord is thicker, firmer, and less vascular than that of the brain.
Spaces of the Meninges
Three spaces located within the meninges are important to note. The epidural or extradural space is a potential space located between the skull and outer layer of the dura layer of the brain. In the vertebral column, the epidural space is between the periosteum and the single dural layer. The subdural space is between the inner dura mater and arachnoid layer. This is a narrow space and is the site of subdural hemorrhage with certain injuries. The third space is the subarachnoid space, which is between the arachnoid and pia mater layers and contains CSF.
CEREBROSPINAL FLUID
CSF is normally a clear, colorless, odorless solution that fills the ventricles of the brain and the subarachnoid space of the brain and spinal cord. The purpose of CSF is to act as a shock absorber, cushioning the brain and spinal cord against injury caused by movement, and to provide nutrients to the neural components. The specific gravity of CSF is 1.007 (see Chapter 6 for CSF normal values). CSF differs from other extracellular fluids in the percentage of composition of various factors. It is composed of water, a small amount of protein, oxygen, and carbon dioxide. The electrolytes—sodium, potassium, and chloride and glucose, an important cerebral nutrient—are also present. An occasional lymphocyte may be present. Normally, CSF pressure is in the range of 60 to 180 mm of water pressure in the lateral recumbent position, which is the position assumed for a lumbar puncture. With the patient in the sitting position, a normal lumbar puncture will register 200 to 350 mm of water pressure. Fluctuation in pressure occurs in response to the cardiac cycle and respirations. The amount of CSF in adults is approximately 125 to 150 mL.
FORMATION OF CEREBROSPINAL FLUID
CSF, which is produced by active transport and diffusion, is formed from three different sources. The major source of CSF is the secretions from the choroid plexus, a cauliflower-like structure located in portions of the lateral, third, and fourth ventricles (Fig. 5-11). The choroid plexus is a collection of blood vessels covered with a thin coating of ependymal cells. CSF is constantly secreted from these surfaces. It is estimated that the amount of CSF produced daily by the choroid plexus is about 500 mL, or 25 mL/hr. A lesser proportion of CSF is secreted from the second source, the ependymal cells, which line the ventricles and blood vessels of the meninges. Finally, CSF is also produced by the blood vessels of the brain and spinal cord. The amount produced from this source is small.
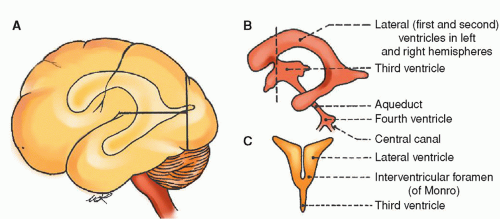
Ventricular System
The two lateral ventricles, one on either side of the midline, are located in the lower and inner parts of the cerebral hemisphere. Each lateral ventricle consists of a central cavity or body and three horns. The central cavity is located in the lower part of the parietal lobe. The anterior horn curves forward and outward into the frontal lobe; the posterior horn curves backward and inward into the occipital lobe; and the middle or lateral horn descends into the temporal lobe. The curved corpus callosum forms the undersurface of the central cavity and the roof of the anterior, middle, and posterior horns.
Figure 5-11 ▪ Diagram of the flow of cerebrospinal fluid from the time of its formation from blood in the choroid plexuses until its return to the blood in the superior sagittal sinus.
The singular third ventricle is a midline inner brain-cavity structure. The two optic thalami form the side walls. The floor is formed by the tuber cinereum (infundibulum, pituitary), corpora albicantia, and crus cerebri. The ventricle is bounded by the fornix in the front and pineal body in the back. The singular, diamondshaped fourth ventricle is laterally bounded by the pons and the superior cerebellar peduncles. The roof is formed by the superior cerebellar peduncles and medulla. The floor of the fourth ventricle is continuous with the central spinal canal.
Flow of Cerebrospinal Fluid
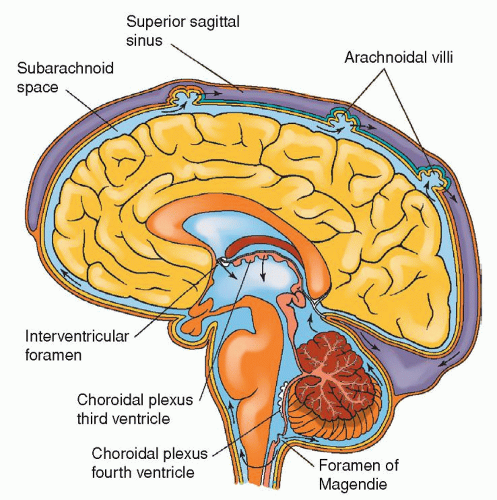
CSF circulation has been termed the “third circulation.” It is a closed system. Fluid formed by choroid plexuses in the two lateral ventricles passes into the third ventricle by way of the two foramina of Monro. The single cerebral aqueduct or aqueduct of Sylvius connects the third and fourth ventricles. CSF flows through the two lateral foramina of Luschka and midline through the foramen of Magendie to the cisternal magnum. At this point, the CSF enters the subarachnoid space. The foramen of Magendie allows CSF to circulate around the cord whereas the foramen of Luschka directs the CSF around the brain (Fig. 5-12).
Figure 5-12 ▪ The cerebral ventricles. A: Lateral aspect of the left cerebral hemisphere showing the contour of the lateral ventricles and their relation to the cerebral lobes. B: Lateral outline of the four ventricles. C: Frontal (coronal) section of the lateral and third ventricles at the level of the dotted line (B), showing their communicating interventricular foramen. (From DeMyer, W. (1998). Neuroanatomy (2nd ed.). Baltimore: Williams & Wilkins.)
Expanded areas of the subarachnoid space are called cisterns. CSF may be aspirated from some of these areas for analysis. The major cisterns are the cisterna magnum, between the medulla and cerebellar region, and the lumbar cistern, between vertebrae L-2 and S-2.
Cerebrospinal Fluid Absorption
Most of the CSF produced daily is reabsorbed into the arachnoid villi, which are projections from the subarachnoid space into the venous sinuses of the brain (Fig. 5-11). CSF drains into the superior sagittal sinus. Arachnoid villi are very permeable and allow CSF, including protein molecules, to exit easily from the subarachnoid space into the venous sinuses. CSF flows in one direction through the arachnoid villi (most of which are located in the subarachnoid space of the cerebrum), which have been compared with pressuresensitive valves. When CSF pressure is greater than venous pressure, CSF leaves the subarachnoid space. As pressures are equalized, the valves close. Dysfunction of CSF flow is seen in hydrocephalus.
CEREBROVASCULAR CIRCULATION
The blood vessels that supply the nervous system form an extensive capillary bed, particularly in the gray matter of the brain. About 20% of the oxygen consumed by the body is used for the oxidation of glucose to provide energy. The brain is totally dependent on glucose for its metabolism. A lack of oxygen to the brain for 5 minutes can result in irreversible brain damage.
The brain receives approximately 750 mL/min of blood, or 15% to 20% of the total resting cardiac output. These figures remain relatively constant because of various control systems affecting the brain. Blood flow rates to specific areas of the brain correlate directly to the metabolism of the cerebral tissue. The brain is supplied by two pairs of arteries: the two internal carotid arteries and the two vertebral arteries. Cerebral circulation is also divided into the anterior and the posterior circulation. The anterior circulation refers to the common carotids and their distal branches including the internal carotid arteries, the middle cerebral arteries, and the anterior cerebral arteries. The posterior circulation refers to the vertebral arteries, the basilar artery, and the posterior arteries.
Internal Carotid Arteries
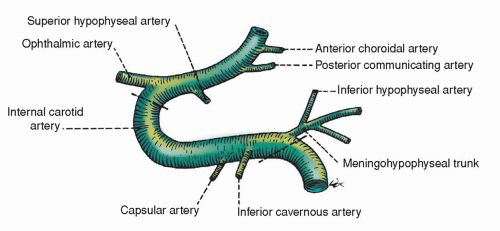
The internal carotid arteries originate from two different vessels: the left common carotid, which originates directly from the aorta, and the right common carotid, which arises from the innominate artery also originating from the aorta. The common carotids branch to form the external and internal carotid arteries. The external carotid artery supplies the face, scalp, and other extracranial structures. The internal carotid artery enters the cranial vault through the foramen lacerum in the floor of the middle cranial fossa. As the internal carotid passes through the bone and dura at the base of the skull and approaches the upper brain, it passes forward through the cavernous sinus just lateral to the pituitary fossa. It curves sharply several times and roughly forms an “S,” called the carotid siphon (Fig. 5-13). Most of the hemispheres, excluding the occipitals, the basal ganglia, and the upper two thirds of the diencephalon, are supplied by the internal carotid arteries (Fig. 5-14).
Figure 5-13 ▪ Lateral view of the left carotid artery at the siphon or parasellar region. The lower interrupted line is where the carotid artery enters the cavernous sinus, and the upper interrupted line is where it exits. (From DeMyer, W. (1998). Neuroanatomy (2nd ed.). Baltimore: Williams & Wilkins.)
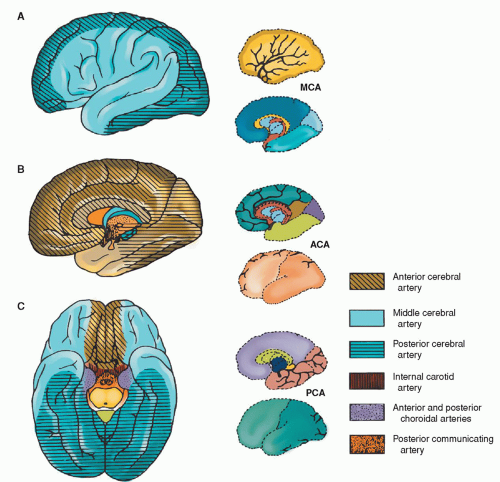
Figure 5-14 ▪ Surface distribution of the anterior, middle, and posterior cerebral arteries. A: Lateral view of the left cerebral hemisphere. B: Medial view of the left cerebral hemisphere. C: Ventral view of the cerebrum. ACA, anterior cerebral artery; MCA, middle cerebral artery; PCA, posterior cerebral artery. (From DeMyer, W. (1998). Neuroanatomy (2nd ed.). Baltimore: Williams & Wilkins.)
TABLE 5-3 MAJOR INTERNAL CAROTID ARTERIAL BRANCHES AND THE CEREBRAL AREAS THEY INNERVATE
ARTERY
AREA SUPPLIED
Ophthalmic
Orbits and optic nerves
Posterior communicating (PCOM)
Connects the carotid circulation with the vertebrobasilar circulation
Anterior choroidal
Part of choroid plexuses of lateral ventricles; hippocampal formation; portions of globus pallidus; part of internal capsule; part of amygdaloid nucleus; part of caudate nucleus; part of putamen
Anterior cerebral (ACA)
Medial surfaces of frontal and parietal lobes; part of cingulate gyrus and “leg area” of precentral gyrus
Recurrent artery of Heubner
Special branch of ACA, penetrates the anterior perforated substance to supply part of basal ganglia and genu of internal capsule (also called medial striate artery)
Middle cerebral (MCA) (has several branches)
Entire lateral surfaces of the hemisphere except for the occipital pole and the inferolateral surface of the hemisphere (supplied by posterior cerebral artery)
Lenticulostriate (from MCA)
Part of basal ganglia and internal capsule
Anterior communicating (ACOM)
Connects the two ACAs
The first intracranial branch from the internal carotid artery is the ophthalmic artery. The terminal branches of each carotid include the posterior communicating artery, the anterior cerebral artery, and the middle cerebral artery. Table 5-3 summarizes the major branches of the carotid arteries and the areas supplied (also see Figure 5-14). See also Chapter 22 for further discussion of cerebral vascular territories and stroke.
CHARACTERISTICS OF THE CEREBRAL CIRCULATION
The following lists the outstanding characteristics of cerebral circulation.
Cerebral arteries have thinner walls than arteries of comparable size in other parts of the body. Cerebral arteries have an internal elastic tissue and scanty smooth muscle.
The veins (other than sinuses) have even thinner walls in proportion to their size and lack a muscle layer. The veins and sinuses have no valves. The venous return does not retrace the course of corresponding arteries but follows a pattern of its own. The dural sinuses are unique to cerebral circulation.
The distribution of the arteries with rich surfaces (arteries anastomosed by localized “end-artery” distribution of branches penetrating the nervous tissue) is distinctive.
Vertebral Arteries
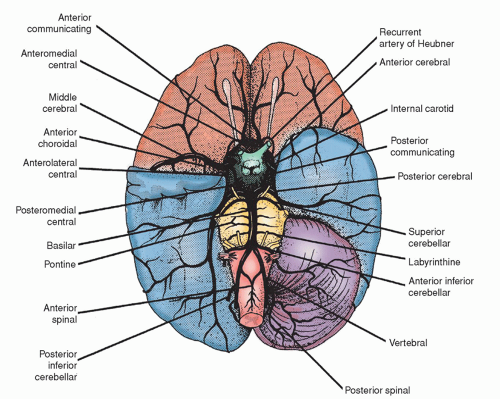
The vertebral arteries, originating from the subclavian arteries, enter the skull through the foramen magnum, ventrolateral to the spinal cord. The two vertebral arteries unite at the level of the pons to become the singular basilar artery (Fig. 5-15). The basilar artery subdivides into the two posterior cerebral arteries that supply part of the cerebrum (Fig. 5-16).
In general, the vertebral arteries and their branches supply the cerebellum, the brainstem, the spinal cord, the occipital lobes, the medial and inferior surfaces of the temporal lobes, and the posterior diencephalon.
Before they begin to supply blood to the brain, the vertebral arteries give off recurrent branches that anastomose with the anterior and posterior spinal arteries and with a posterior meningeal branch. In its intracranial course, the vertebral arteries give rise to direct bulbar arteries to the medulla, the anterior spinal artery, the posterior inferior cerebellar artery (PICA), sometimes the posterior spinal artery, and small branches to the basal meninges. The first branch of the basilar artery is the anterior inferior cerebellar artery (AICA). The basilar artery is also the origin of the pontine arteries, the internal auditory arteries, the superior cerebellar arteries, and the posterior cerebral arteries. Table 5-4 summarizes the major branches of the vertebral arteries and the areas supplied.
Circle of Willis
The circle of Willis, which is located at the base of the skull, is divided into anterior (carotid portion) and posterior (vertebrobasilar portion) circulation (Fig. 5-15). The composition of each portion includes these elements:
TABLE 5-4 MAJOR VERTEBRAL ARTERIAL BRANCHES AND THE CEREBRAL AREAS THEY INNERVATE
ARTERY
AREA SUPPLIED
Vertebral Branches
Anterior spinal (only one artery)
Anterior two thirds of spinal cord
Posterior spinals
Posterior one third of spinal cord
Posterior inferior cerebellar (PICA)
Undersurface of the cerebellum, medulla, and choroid plexuses of fourth ventricle
Basilar Artery Branches
Posterior cerebral (PCA)
Occipital lobes, medial and inferior surfaces of the temporal lobes, midbrain, and choroid plexuses of third and lateral ventricle
Posterior choroidals (from PCA)
Medial posterior choroidal
Lateral posterior choroidal
Tectum, choroid plexus of third ventricle, and superior and medial surfaces of the thalamus
Penetrating the choroidal fissure and anastomosing with branches of the anterior choroidal arteries
Anterior inferior cerebellar (AICA)
Undersurface of the cerebellum and lateral surface of the pons
Superior cerebellar (SCA)
Upper surface of the cerebellum and midbrain
Pontine
Pons
Figure 5-15 ▪ The blood supply of the brain, as seen on the ventral surface. (The right cerebellar hemisphere and the tip of the right temporal lobe have been removed.) (From Barr, M. L., & Kiernan, J. A. (1993). The human nervous system (6th ed.). Philadelphia: J. B. Lippincott.)
Middle cerebral arteries, the anterior cerebral arteries, and the anterior communicating artery, which connects the two anterior cerebral arteries.
Two posterior cerebral arteries; two posterior communicating arteries connect the middle cerebral arteries with the posterior cerebral arteries, thus uniting the internal carotid system with the vertebral-basilar system.
The circle of Willis encloses a very small area that is little more than 1 in2 (6.45 cm2), or about the size of a quarter. Functionally, the carotid circulation and the posterior circulation usually remain separate. At one time, the circle of Willis was thought to be a protective mechanism by which blood was shunted to compensate for alterations in cerebral blood flow or pressure. However, collateral circulation through the circle depends on the patency of its components. The vessels of the circle of Willis, particularly the communicating arteries, may be anomalous. Nevertheless, in favorable instances, the circle does permit an adequate blood supply to reach all parts of the brain, even after one or more of the four supplying vessels has been ligated.
Only gold members can continue reading. Log In or Register to continue